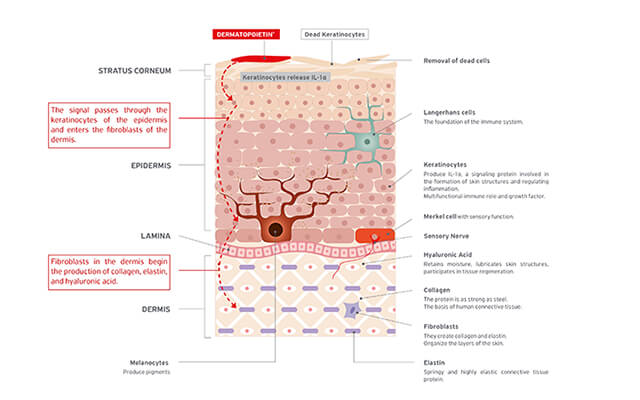
Wir bei EVENSWISS sind überzeugt, dass jedermann und jedefrau gesunde und jugendlich aussehende Haut und Haare verdienen. Mit diesem Ziel vor Augen verfolgen EVENSWISS® Produkte einen revolutionären Ansatz in der Hautpflege, indem sie auf der Oberfläche der Haut eine biologische Signalkette hervorrufen, die tief in die Haut eindringt und deren Erneuerung auslöst: signalling cosmetics. Diesem Ziel verpflichtet, unternehmen wir alles, um fortschrittliche und wirksame Pflegeprodukte zu entwickeln, die in einem tiefen Verständnis der biologischen Mechanismen der Erneuerung von Haut und Haar gründen. Basierend auf den Erkenntnissen von 30 Jahre Forschung konnten wir eine weltweit einzigartige Formel entwickeln: EVENSWISS® with Dermatopoietin®.
26 Dezember 2023